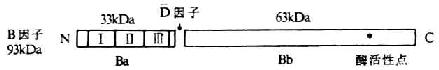
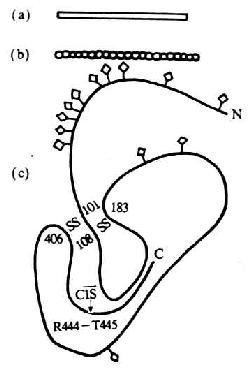
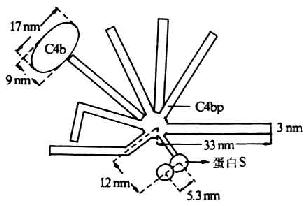
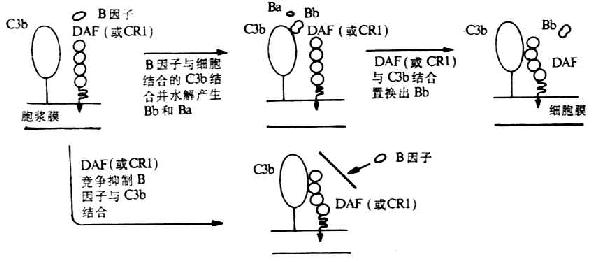
第一章 白细胞分化抗原
机体免疫系统是由中枢淋巴器官、外周淋巴器官、免疫细胞和免疫分子所组成。免疫应答过程有赖于免疫系统中细胞间的相互作用,包括细胞间直接接触和通过释放细胞因子或其它介质的相互作用。免疫细胞间或细胞与介质间相互识别的物质基础是免疫细胞膜分子,包括细胞表面的多种抗原、受体和其它分子。细胞膜分子通常也称为细胞表面标记(cell surface marker)。免疫细胞膜分子的研究对于深入了解免疫应答的本质以及临床某些疾病的诊断、预防和治疗都具有十分重要的意义。
免疫细胞膜分子的种类相当繁多,主要有T细胞受体,B细胞识别抗原的膜免疫球蛋白,主要组织相容性复合体抗原,白细胞分化抗原,粘附分子,结合促分裂素的分子,细胞因子受体,免疫球蛋白Fc段受体以及其它受体和分子。白细胞分化抗原是白细胞(还包括血小板、血管内皮细胞等)在分化成熟为不同谱系(lin-eage)和分化不同阶段以及活化过程中,出现或消失的细胞表面标记。它们大都是穿膜的蛋白或糖蛋白,含胞膜外区、穿膜区和胞浆区;有些白细胞分化抗原是以糖基磷脂酰肌醇(glyco-sylphosphatidylinositol,GPI)连接方式“锚”在细胞膜上。少数白细胞分化抗原是碳水化合物半抗原。白细胞分化抗原参与机体重要的生理和病理过程。假如:(1)免疫应答过程中免疫细胞的相互识别,免疫细胞抗原识别、活化、增殖和分化,免疫效应功能的发挥;(2)造血细胞的分化和造血过程的调控;(3)炎症发生;(4)细胞的迁移如肿瘤细胞的转移等。
第一节 白细胞分化抗原的分类
80年代初以来,由于单克隆抗体,分子克隆、基因转染细胞系等技术在白细胞分化抗原研究中得到广泛深入的应用,有关白细胞分化抗原的研究和应用进展相当迅速。在世界卫生组织(WHO)和国际免疫学会联合会(IUIS)的组织下,自1982年至1993年已先后举行了五次有关人类白细胞分化抗原的国际协作组会议(International workshop on human leukocyte differentiation antigens),并应用以单克隆抗体鉴定为主的聚类分析法,将识别同一分化抗原的来自不同实验室的单克隆抗体归为一个分化群,简称CD(cluster of differentiation)。在许多场合下,抗体及其识别的相应抗原都用同一个CD序号,因此在参阅教科书和文献时需加注意。
一、人白细胞分化抗原的分类
迄今为至,人CD的序号已从CD1命名至CDw130(见表1-2),可大致划分为T细胞、B细胞、髓系细胞、NK细胞、血小板、激活抗原、粘附分子、内皮细胞和细胞因子受体等九个组(表1-1)。
表1-1 CD单抗分组(1993)
| 主要特异性 | CD |
| T细胞 | CD1-CD8、CD27、CD28、CD38、CD39、CDw60、CD45、CD45RA、CD45RB、CD45RO、CD98、CD99、CD99R、CD100、CDw101 |
| B细胞 | CD10、CD19-CD24、CD37、CD40、CD53、CD72-CD75、CDw76、CD77、CD78、CD79a、CD79b、CD80-CD83、CDw84、CD85、CD86 |
| 髓系细胞 | CDw12、CD13-CDw17、CD32-CD35、CD64、CDw65、CD66a-CD68、CD87-CD93 |
| NK细胞 | CD56、CD57、CD94 |
| 血小板 | CD9、CD31、CD36、CD4la、CD4lb、CD42a-CD42d、CD61、CD63、CD107a、CD107b、CD26、CD30、CD69、CD70、CD71、CD95-CD97 |
| 激活抗原 | CD26、CD30、CD69、CD70、CD71、CD95-CD97 |
| 粘附分子 | CD11a-CD11c、CD15s、CD18、CD29、CD43、CD44、CD44R、CD48、CD49a-CD49f、CD50、CD51/CD61、CD54、CD55、CD58、CD59、CD62E、CD62L、CD62P、CD102-CD104、CDw108 |
| 内皮细胞 | CD105、CD106、CDw109 |
| 细胞因子受体 | CD25、CD115、CDw116、CD117、CDw119、CD120a、CD120b、CDw121a、CDw121b、CD122、CDw124、CD126、CDw127、CDw128、CDw130 |
注:(1)CD是流水编号,但CD110~CD114,CD118、CD123、CD125和CD129暂缺;CD67和CD66b是重复的。
(2)凡CD中带有w的抗原或抗体发CDw108、CDw109尚需继续进行全面鉴定。
(3)有些CD抗原又可进一步划分为不同的成员,一般用小写英文字母表示,但情况有所不同:1)如CD1可分为CD1a、CD1b和CD1c,这三种不同分子是分别由三个不同的、高度同源的基因所编码;2)CD45至少可分为CD45R、CD45RA、CD45RB和CD45RO,它们是同一基因的不同异型(isoform);3)CD2和CD2R是识别同一个分子上不同的表位;4)CD49已进一步划分为CD49a、CD49b、CD49c、CD49d、CD49e和CD49f,它们的基因定位于不同的染色体上,但具有较高的同源性。
(4)CD九个组划分的特异性是相对的,实际上,许多CD抗原的组织细胞分布较为广泛。此外,有的CD抗原可由不同的分类角度而归入不同的组,如IL-2受体a链CD25是活化T细胞的标记,也属于细胞因子受体;再如某些属于T细胞、B细胞、髓系细胞或NK细胞组的CD抗原实际上也是粘附分子。
表1-2 CD分子的主要特征
| CD | 常用单克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CDla | T6,Leu6 | Thy,DC,LHC | gp49 | TCRγδ的配体? |
| CDlb | WM-25,4A76,NUT2 | Thy,LHC | gp45 | TCRγδ的配体? |
| CDlc | L161,M241,7C6 | Thy,DC,Bsub | gp43 | TCRγδ的配体? |
| CD2 | 9.6,T11,Leu5;(LFA-2,SRBC-R) | T,NKsub. | gp50 | 与LFA-3(CD58)和CD59结合,T细胞活化 |
| CD2R | T11.3,9.1 | Ta | gp50 | T细胞活化 |
| CD3 | T3,Leu4 | T | γ、δ、ε、ζ、η5种链分别为p26,20,19,16,21 | T细胞活化 |
| CD4 | T4,Leu3a | Tsub | gp55 | 与MHCⅡ类分子结合,信号转导,HIV受体 |
| CD5 | T1,UCHT2,T101,Leu1 | T,Bsub | gp67(小鼠Lyt-1类同物) | 与CD72结合,T细胞信号转导 |
| CD6 | T12,T411 | Tsub,Bsub | gp100 | ? |
| CD7 | 3A1,Leu9 | Tsub,(NK,Pt) | gp40 | ? |
| CD8 | α链:T8,Leu2a,UCHT4 β链:T8/2T8,5H7 | Tsub(αβ),NKsub(α/α) | gp(36/32),α/α或α/β二聚体 | 与MHCI类分子结合,信号转导 |
| CD9 | PHN200,FMC56 | Pre-B,M,Pt | p24(TM-4成员) | 血小板活化 |
| CD10 | J5;(CALLA) | Pre-B,CALlG | gp100 | 中性肽链内切酶,水解脑啡肽、趋化肽和P物质 |
| CD11a | MHM24,2F12;CRIS-3 | Leu | LFA-1(gp180/95)的α链 | 与ICAM-1(CD54)和ICAM-2(CD102)结合 |
| CD11b | Mol,OKM1;(Mac-1,CR3,integrin αm | G,M,NK | gp165/95的α链 | C3bi和FB受体,与ICAM-1结合,粘附,调理吞噬 |
| CD11c | LeuM5;(CR4,integrin αx) | M,G,NK,Tsub | gp150/95的α链 | C3bi受体,调理吞噬 |
| CDw12 | M67 | M,G,(Pt) | p90~120 | ? |
| CD13 | My7, MOU28 | M,G | gp150 | 氨肽酶 |
| CD14 | Mo2,UCHM1,LeuM3 | M,(G,LHC) | gp55(GPI连接) | LPS/LPS结合蛋白复合体受体 |
| CD15 | MY1,LeuM1 | G,(M),RS | Lewis*3FAL,X-hapten,Lex | 参与吞噬 |
| CD15s | (Sialyl CD15) | G,M | SialylLewisx(sLex) | CD62E和CD62P配体,白细胞粘附到En和Pt |
| CD16a | HUNK2,Leu11,MEM-154(FcγRⅢA/FcγRⅢB) | NK,G,Mo,Mac | gp50-70(穿膜形式) | ADCC,NK活化 |
| CD16b | ID3(FcγRⅢB) | G,M | 48(GPI连接) | ? |
| CDw17 | GO35 | G,M,Pt | 乳糖基酰鞘氨醇 | ? |
| CD18 | MHM23;(LFA组β链,integrin β2) | Leu | gp95,LFA-1,CR3、p150/95的β连 | ICAM-1(CD54)、ICAM-2(CD102)、C3bi配体,粘附,调理吞噬 |
| CD19 | B4,Leu12 | B | gp90 | 调节B细胞活化 |
| CD20 | B1,Leu16 | B | p37/35(非糖基化穿膜磷蛋白) | Ca2+通道?调节B细胞活化 |
| CD21 | B2,OKB-1;(CR2) | Bm,FDC | p140 | C3d/EBV受体,B细胞活化,结合sCD23 |
| CD22 | HD39,Leu14,SHCL-1,HC2 BL-CAM | B | gp130/140,髓鞘(磷)脂相关蛋白类似物(MAG) | 与CD45RO结合,B细胞粘附、B细胞活化? |
| CD23 | MHM6,Leu20;(FcεRⅡ) | Bm,Ba,,Ma,Eo | gp45~50 | 参与IgE生成的调节,调节B细胞分化,IgE介导的ADCC,结合CD21? |
| CD24 | BA-1 | B,G | gp41/38 | ? |
| CD25 | TAC,7G7/B6;(IL-2Rα) | Ta,Ba,Ma | gp55 | 组成高亲力受体,T细胞生长 |
| CD26 | 5.9,Tal | Ta,B,Mac | gp120 | 二肽酰肽酶Ⅳ(DPPⅣ),HIV另一类受体? |
| CD27 | VIT14,S152,OKT18A | Tsub | p55,NGF受体家庭 | CD70的配体 |
| CD28 | 9.3 | Tsub,Ba | gp44 | 与CD80、CD86互为配体 |
续表2
| CD | 常用克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CD29 | 4B4,(integrinβ1,FNRβ) | 广泛分布 | gp130,GPⅡa | 与ECM粘合,细胞间粘附,结合VCAM-1(CD106) |
| CD30 | Ki-1 | Ta,Ba,Rs | gp105,NGF受体家族 | 与淋巴细胞存活和增殖有关 |
| CD31 | SG134,TM3,HEC-75;(PECAM) | Pt,M,G,B | gp140,血小板GPⅡa | 粘附 |
| CD32 | CIKM5,41H16(FcγRⅡ) | Mac,G,B,Eo | gp40 | 凝聚IgG FcR,吞噬,ADCC |
| CD33 | MY9,H153,L4F3 | M,BM | gp67 | ? |
| CD34 | MY10,ICH3 | BM | gp105~120 | 生长因子受体?调控早期造血 |
| CD35 | TO5,E11,(CR1) | G,M,B,NKsub,RBC | p160~260 | 结合C3b,调理吞噬 |
| CD36 | 5E1,ESIVC7,OKM5 | M,Pt,(B) | gp88,血小板GPⅢb | 结合ECM,血小板粘附 |
| CD37 | HD28,HH1 | Bm,(T,M) | gp40~52 | ? |
| CD38 | Leu17,T16,OKT10 | PC,Ta,Thy, | p45 | ? |
| CD39 | AC2,G28-10 | Bm,(M),FDC | gp70~100 | ? |
| CD40 | G28-5,EA-5 | B,FDC | p50,NGF受体家族 | B细胞生长和记忆细胞产生 |
| CD41 | PBM6.4,PL273;(integrinαⅡb) | Pt | GPⅡb/Ⅲa中的GPⅡb(gp120/25) | 血小板凝集和活化Fb,结合ECM的受体 |
| CD42a | FMC25,GR-P | Pt,Meg | gp23,血小板 GPⅨ,形成GPIb/Ⅸ复合物 | 血小板粘附,结合vWF |
| CD42b | PHN89,AN51 | Pt,Meg | gp135/25,血小板GP1b-α形成GPIb/Ⅸ复合物 | 血小板粘附,结合vWF |
| CD42c | Pt,Meg | 22,血小板GPIb-β | ||
| CD42d | Pt,Meg | 85,血小板GPV | ||
| CD43 | OTH71C5,G19-1;(Leukosialin) | T,G,M | gp95,Sialophorin | T细胞活化?与CD54结合 |
续表3
| CD | 常用况隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CD44 | GRHL1,Hermes,F10-44-2(Pgp-1,ECM-RⅢ) | Leu | gp80~215 | 粘附ECM,T细胞活化,淋巴细胞归位受体 |
| CD44R | FW11,24 | CD44限制性表位(外显子v9剪接的变异体) | ||
| CD45 | T29/33,BMAC1;(T200) | Leu | p170~240,白细胞共同抗原(LCA) | PTP酶,调节信号传导调节信号传导 |
| CD45RA | G1-15,F8-11-13,Leu18;(限制性LCA) | Tsub,B,M,(G,NK) | gp220 | 调节信号传导 |
| CD45RB | PT17/26/16;(限制性LCA) | Leu | gp190/205/220 | 调节信号传导 |
| CD45RO | UCHL1;(限制性LCA) | Tsub,Bsub,G,M | gp190 | 与CD22结合,调节信号传导 |
| CD46 | HULYM5,J48 | Leu,Pt | 膜辅助因子蛋白(MCP),gp56/66 | 调节补体活化,麻疹病毒受体 |
| CD47 | BRIC126,CIKM1 | 广泛分布 | gp47~52,N连接葡聚糖 | ? |
| CD48 | WM68,LO-MN25 | Leu | gp41(GPI联结),与CD58有68%同原 | CD2的配体 |
| CD49a | SR84,IB3.1(VLAα1) | T,M | gp210,与CD29组成VLA-1 | 粘附CA和LM |
| CD49b | Gil4(VLA-α2,ECMR-Ⅱ,Pt-GPIa) | Leu,Pt | gp165,与CD29组成VLA-2 | 粘附CA |
| CD49c | J143(VLAα3,ECMR-1) | T,Bsub,M | gp135/25,与CD29组成VLA-3 | 粘附FN、CA和 LM |
| CD49d | B5G10,HP2/1;(VLA-α4) | M,T,B,Thy,Pt | gp150,80,70,与CD29组成VLA-4 | 粘附FN,结合VCAM-1(CD106),归位受体 |
| CD49e | 2H6,3D3(VLAα5,FNRα,ECMR-Ⅳ) | T,Bsub,m | gp130/25,与CD29组成VLA-5 | 粘附FN |
| CD49f | GOH3(VLA-α6) | Pt,(T) | gp120/30,与CD29组成VLA-6 | 粘附LM |
| CD50 | 101-1D2,140-11,(ICAM-3) | Leu | gp140/108 | 粘附,CD11a-CD11b/CD18)配基 |
| CD51 | 13C2;23C6;NK1-M7(VNRα链,integrin αv) | Pt,Leu | gp125/24,与CD61组成二聚体 | 粘附VN、FN和vWF |
续表4
| CD | 常用克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CD52 | YTH66.9;(Campath-1) | Leu | gp21~28 | ? |
| CD53 | HI36,MEM-53,HD77 | leu,BM | gp32~40(TM-4成员) | ? |
| CD54 | WEHI-CAMI,OKT27(ICAM-1) | 广泛分布 | gp90(80~114),细胞间粘附分子-1 | 与LFA-1和CD43结合,鼻病毒受体,En上恶性疟原虫受体 |
| CD55 | 143-30,BRIC110,BRIC123;(DAF) | 广泛分布 | p70,衰变加速因子(GPI联结) | 调节补体活化 |
| CD56 | Leu19,NKH1;(N-CAM) | NK,(Tsub) | 神经细胞粘附分子(N-CAM)的三种异构体gp120,140,180 | 粘附 |
| CD57 | Leu7,HNK-1 | NK,Tsub,Bsub | gp110 | ? |
| CD58 | G26,BRIC5;(LFA-3) | 广泛分布 | 白细胞功能抗原-3,gp40~65(部分GPI联结) | 与CD2结合,粘附 |
| CD59 | MEM-43,YTH53.1;(TAP,Protectin) | 广泛分布 | gp18~20(GPI联结) | 与CD2结合,抑制MAC |
| CDw60 | M-T32,M-T21,M-T41;(GD3) | Tsub,Pt | 乙酰神经氨酸-乙酰神经氨酸半乳糖p105 | ? |
| CD61 | Y2/51,CLB-thromb/1(VNR-β链, integrinβ3) | Pt,Meg | p105血小板GPⅢa,与CD51组成VNR | 结合VN、FN和vWF |
| CD62E | 3B7,4DIO(E-selectin,ELAM-1) | En | gp115 | 粘附L-selectin、CD15s |
| CD62L | Leu8,FMC46(L-selectin,LAM-1) | T,B,M,NK | gp75~80 | 粘附E-selectin,P-selectin? |
| CD62P | G2,AK-6(P-selectin,GMP-140,PADGEM) | Pt,En | gp140,Lectin family血小板α颗粒 | 结合PMN、M表面 |
| CD63 | RUU-SP2.28,CLB | Pt,M,Mac,(G,T,B) | gp53,血小板致密颗粒(TM-4成员) | CD15s,粘附到En和Pt? |
续表5
| CD | 常用克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CD64 | MAb32.2,MAb22;(FcγRI) | M | gp70 | 吞噬、ADCC, Mac活化 |
| CDw65 | VIM2,HE10,CF4 | G,M | 岩藻糖基神经节苷脂 | 中性粒细胞活化 |
| CD66a | BGP | 髓样细胞 | 180~200(胆汁糖蛋白-1) | ? |
| CD66b | MF25·1(P100,原CD67) | G | 95~100 | ? |
| CD66c | NCA | 髓样细胞 | 90~95 | ? |
| CD66d | CGM1 | 髓样细胞 | 30 | ? |
| CD66e | CLB-gran/10(CEA) | 髓样细胞,上皮 | gp180~200 | 粘附 |
| CD67 | (CD66b) | G | 95~100 | ? |
| CD68 | EBM11,Ki-M7,Ki-M6 | Mac | gp110 | ? |
| CD69 | Leu23(AIM) | Ta,Ba,Mac,NK | 34,28 | 活化诱导分子 |
| CD70 | Ki-24(CD27L) | Ta,Ba,RS | 55,75,95,110,170 | CD27的配体 |
| CD71 | OKT9;(TfR) | Mac,增殖细胞 | p95 | 铁代谢,细胞生长 |
| CD72 | S-HCL2,J3-109,BU-40 | B | gp43,39 | 与CD5结合 |
| CD73 | 7G2.2.11,AD-2 | Bsub,Tsub | p69(GPI连结) | 5`一核苷酸外切酶 |
| CD74 | LN2,BU-43 | B,M | gp41/35/33.Ⅱ类相关恒定链(γ链) | 与新合成MHCⅡ类分子结合,参与抗原提呈 |
| CDw75 | LN1,HH2 | Bm,Tsub | p53,α2.6sialyltransferase | 酶活性 |
| CDw76 | HD66,CRIS-4 | Bm,Tsub | p85/67 | ? |
| CD77 | 38.13(BLA);424/4A11 | B | Globotriaosylceramide(Gb3) | ? |
| CDw78 | Leu21,AntiBa | B | p67? | ? |
| CD79a | mb-1(Igα) | B | 33,40 | mIg(BCR)复合成分 |
| cd79b | B29(Igβ) | B | 33,40 | mIg(BCR)复合 |
| CD80 | B7-1,BB1 | Ba,Mac,胸腺Stromal cell | 60 | 活化B细胞抗原,CD28、CTLA-4配体,刺激T细胞活化 |
续表6.
| CD | 常用克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CD81 | ID6,5A6(TAPA-1) | B,T,M | 22(TM-4成员) | 离子通道,增殖抗体靶抗原 |
| CD82 | R2,1A4,4F9 | B | 50~53(TM-4) | 信号传递 |
| CD83 | HB15 | B,DC | 43 | ? |
| CDw84 | 2G7,152-ID5 | B | 73 | ? |
| CD85 | VWP-55,GH1/75 | B,M,PC | 120,83 | ? |
| CD86 | FUN-1,BU63 | Bac,Tac,M | 80 | CD28配体,活化T细胞信号 |
| CD87 | UPA-R | 髓样细胞 | 50~65(GPI连接) | 结合尿激酶血纤维蛋白溶酶原激活因子 |
| CD88 | S5/1,W17/1(C5aR) | 髓样细胞 | 42 | 补体C5a受体,趋化作用 |
| CD89 | 79E6,A3(FcαR) | 髓样细胞,Tsub,Bsub. | 55~75 | IgA Fc段受体 |
| CDw90 | 5E10(Thy-1) | 髓样细胞,造血祖细胞 | 25~35(GPI连接) | T细胞活化,神经细胞粘附 |
| CD91 | A11,C2(α2M-R) | 髓样细胞 | 517/85(α/β二聚体) | α2-巨球蛋白受体,与M,Mac分化有关 |
| CDw92 | VIM15 | 髓样细胞 | 70 | ? |
| CD93 | VIMD2 | 髓样细胞 | 120 | ? |
| CD94 | HP3Bi(KP43) | NK | 43/43(同源二聚体) | 调节细胞粘附和溶细胞活性 |
| CD95 | 71CC,anti-Fas (APO-1/FAS) | 活化,广泛 | 42(三个富含半胱氨酸重复结构) | 抗CD95McAb可诱导程序性细胞死亡 |
| CD96 | G8.5,TH-111 TACTILE | 活化 | 160 | T细胞活化? |
续表7
| CD | 常用克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CD97 | VIM3b,VIM3C(BL-KDD/F12) | 活化 | 74,80,89 | T细胞活化? |
| CD98 | 4F2,2F3 | Ta,Thy,NK,M,Pt | 80/40(异二聚体) | 激酶相关 |
| CD99 | D44,FMC29(E2,MIC2) | T,Leu | 32 | E花结形成;粘附作用 |
| CD99R | HI170,IT4,(E2,MIC2) | T | 32 | ? |
| CD100 | BD16,BB18,A8 | T,Ta,NK,M | 150 | 细胞激活和增殖 |
| CDw101 | BB27,BA27 | T | 140 | 与CD28共表达 |
| CD102 | CBR-IC2/1,(ICAM-2) | 粘附,Leu,Pt,En | 60 | 粘附,配体LFA-1 |
| CD103 | LF61(HML-1,integrin αE) | 粘附,Tsub | 150,25 | 粘附,T细胞与上皮细胞相互作用 |
| CD104 | 439-9B(β4 integrin) | 粘附,上皮,Thy,En,角朊 | 220(形成α6β4二聚体) | 与细胞骨架相连 |
| CD105 | 44G4,1G2(Endoglin,TGF-βRⅢ) | En,Mac | 95(S-和L-Endoglin不同胞浆区都含RGD) | TGF-βRⅢ,细胞粘附 |
| CD106 | 1G11(VCAM-1,INCAM-110) | En,M,BM | 100,110 | VLA-4配体,参与粘附 |
| CD107a | H5g11,(LAMP-1) | Pt | 110 | 溶酶体相关膜蛋白 |
| CD107b | H4B4(LAMP-2) | Pt | 120 | 血小板激活 |
| CDw108 | MEM-150,MEM-121 | 粘附,Tac | 80(GPI连接) | 细胞活化 |
| CDw109 | 8A3,7D1 | En,Tac,Pt | 170/150(GPI连接) | 细胞活化、增殖和信号传递 |
| CD115 | MR18(CSF-1R,M-CSFR) | 髓样细胞(M,Mac)定向BM | 150(c-fms基因产物) | M-CSF受体,细胞生长和信号传递 |
| CDw116 | DF2714(GM-CSFRα链) | 髓样细胞(PMN,Eo,M,Mac)BM | 75~85(与β链组成高亲和力受体) | GM-CSF受体,细胞生长和分化 |
| CD117 | 17F11(SCFR,cKIT) | Mas,BM | 145 | SCF受体,肥大细胞生长,增强其它细胞因子信号传递 |
续表8
| CD | 常用克隆抗体或代号( ) | 主要表达细胞 | 分子量(kDa)和结构 | 功能 |
| CDw119 | 3B1,B8(IFN-γR) | 广泛 | 90 | IFN-γ受体,细胞活化,MHC抗原表达 |
| CD120a | MR-1(TNFR;55kD) | 广泛 | 55,NGF受体家族 | TNF受体,与Fas抗原共调变 |
| CDw120b | MR2-1(TNFR;75kD) | T,B,M | 75,NGF受体家族 | TNF受体 |
| CDw121a | hIL-1R1-M1(IL-1RⅠ型) | 广泛,T,B,En,Eb,上皮 | 80 | IL-1受体 |
| CDw121b | hIL-1R2-M22(IL-1RⅡ型) | 广泛 | 68 | IL-1受体,T细胞活化 |
| CD122 | 2RB(IL-2R,75kD,IL-2Rβ) | T,B,NK | 75 | IL-2受体,激活T、B和M |
| CDw124 | hIL-4R-M57(IL-4R) | 造血细胞,Fb,上皮 | 140 | IL-4受体,细胞生长、分化 |
| CD126 | B-C22(IL-6R) | T,Bac | 80(与gp130组成高亲和力受体) | IL-6受体,细胞生长、分化 |
| CDw127 | H2,hIL-7R-M20(IL-7R) | 淋巴样,髓样细胞 | 75 | IL-7受体,细胞生长 |
| CDw128 | GB20(IL-8R) | PMN,Eo,B,M | 58~67 | IL-8受体,趋化和活化PMN |
| CDw130 | AM64(IL-6R-gp130SIG) | 广泛 | 130(与IL-6R,IL-11R,LIFR、OSMR和CNTFR组成高亲和力受体) | IL-6受体gp130,转导信号 |
注:Thy:胸腺细胞;DC:树突状细胞:FDC:滤泡树突状细胞;B:B细胞;Bsub:B亚群;Pre-B,前B细胞;Bm:成熟B细胞;Ba:活化B细胞;T:T细胞;Tsub:T亚群;Ta:活化T细胞;M:Ma:活化单核细胞;Mac:巨噬细胞;Mas:肥大细胞;PC:浆细胞;G:粒细胞;PMN:多形核细胞;My:髓样细胞;NK:自然杀伤细胞;NKsub:NK亚群;LHC:表皮郎罕氏细胞;RS:Reed-Srtenterg细胞;NEC:神经内分泌细胞;RBC:细细胞;Pt:血小板;Eo:嗜酸性粒细胞;BM:骨髓细胞;Meg:巨核细胞;Fb:成纤维细胞;En:内皮细胞;Leu:白细胞;gp:糖蛋白;p:蛋白质;VLA:很晚出现在抗原;CALLA:共同型急性淋巴母细胞白血病抗原;LAMP:溶酶体相关膜蛋白;MCP:膜辅蛋白;MAC:膜攻击单位;LFA:淋巴细胞功能相关抗原;CR:补体受体;3FAL:3-fucosyl-N-acetyl-lactosamine;PECAM:血小板内皮细胞粘附分子;ECMR:细胞外基质受体;LCA:淋巴细胞共同抗原;PTPase:磷酸酪氨酸磷酸酯酶;ICAM:细胞间粘附分子;N-CAM:神经细胞粘附分子;TAP:T细胞活化蛋白;Tyr-P:磷酸化酪氨酸;VCAM:血管细胞粘附分子;GPI:糖基磷脂酰肌醇;AIM:活化诱导分子;LIF:白血病抑制因子;OSM:抑瘤素-M;CNTF:睫状神经营养因子;CA:胶原蛋白;LM:层粘连蛋白;FN:纤粘连蛋白;FB:血纤维蛋白原;vWF:von Willbrand因子;TM-4:四次跨膜家族。
二、小鼠白细胞分化抗原
大多数白细胞分化抗原在生物进化过程中具有保守性,这是不同种的动物执行相同或相似生物学功能的需要。小鼠是免疫学常用的实验动物,而且对某些人白细胞分化抗原的结构和功能的了解首先是从小鼠或小鼠源性的细胞实验模型得知的,表1-3列举了部分与人CD抗原类同的小鼠造血细胞表面抗原,以供参考。
表1-3 与人CD抗原类同的小鼠造血细胞表面抗原
| 小鼠表面抗原 | CD类同物 | 分 布 | 功 能 | 分子量(kDa) | 染色体定位 |
| Lyt-1 | CD5 | T,B亚群 | 70 | 19 | |
| Lyt-2 | CD8a | CTL | CTL粘附 | 30 | 6 |
| Lyt-3 | CD8b | CTL | CTL粘附 | 35 | 6 |
| L3T4 | CD4 | Th/Ti | 结合MHCⅡ类分子 | 52 | 6 |
| Ly-5 | CD45 | 白细胞,干细胞,滤泡状树突细胞,有核红细胞,胸腺细胞 | B细胞成熟 | 200,210,200,190 | 1 |
| Ly-5 | CD45R? | 前B,B,CTL亚群 | 220 | 1 | |
| Ly-15 | CD11a | T,B,髓样细胞,NK,红样细胞,髓样干细胞 | CTL粘附 | 177 | 7 |
| Ly-17 | CD32 | B,髓样,干细胞,T?郎格罕细胞? | IgG2b/1Fc受体 | 55~60 | 1 |
| Ly-37 | CD2 | T,B,Thy | T细胞活化,红细胞受体 | 50~60 | 3 |
| Ly-38 | CD1 | 3 | |||
| Ly-40 | CD11b | Mδ,B,Lyt-1阳性B细胞 | C3bi受体 | 165 | |
| Ly-43 | CD23 | B | IgE Fc受体 | 49 | 1 |
| Ly-42 | CD25 | T,B | IL-2受体α链 | 47~53 | |
| Ly-44 | CD20 | B |
第二节 白细胞分化抗原的应用
CD抗原及其相应的单克隆抗体在基础和临床免疫学研究中已得到广泛的应用。在基础免疫学研究中,CD主要应用于:(1)CD抗原的基因克隆,新CD抗原及新配体的发现;(2)CD抗原结构与功能关系;(3)细胞激活途径和膜信号的传导;(4)细胞分化过程中的调控;(5)细胞亚群的功能。在临床免疫学研究中,CD单克隆抗体可用于:(1)机体免疫功能的检测;(2)白血病、淋巴瘤免疫分型;(3)免疫毒素用于肿瘤治疗、骨髓移植以及移植排斥反应的防治;(4)体内免疫调节治疗。有关与免疫功能相关的CD分子归纳于表1-4。有关与T细胞表面分子、B细胞表面分子以及NK细胞的表面标记参见第七章。与CD有关的Ig超家族、粘附分子、补体受体、细胞因子受体等分别在本书的有关章节中加以介绍。
表1-4 与免疫功能有关的CD
| 免疫功能 | CD |
| 细胞受体 TCR CD3、CD4、CD8mIg mb-1/Igα(CD79a)、B29/Igβ/(CD79b)CR CR1(CD35)、CR2(CD21)、CD3(CD11b/CD18)、CR4(CD11c/CD18)、C5aR(CD88) FcR FcγRI(CD64)、FcγRⅡ(CD32)、FcγRⅢ(CD16)、FcεRⅡ(CD23)FcαR(CD89) 细胞因子受体IL-2Rα(CD25)、M-CSFR(CD115)、GM-CSFR(CDw116)、SCFR(CD117)、 IFN-γR(CDw119)、TNF-αR(CD120)、IL-1R(CDw121)、Il -2Rβ(CD122)、IL-4R(CDw124)、IL-6R(CD126)、IL-7R(CDw127)、IL-8R(CDw128)、gp130(CDw130)细胞间、细胞基质相互识别白细胞粘附分子-内皮细胞粘附分子:LFA-1(CD11a/CD18)-ICAM-1(CD54)、ICAM-2(CD102)Mac-1(CD11b/CD18)-ICAM-1(CD54)VLA-4(CD49d/CD29)-VACM-1(CD106)L-selectin(CD62L)-E-selectin(CD62E)、P-selectin(CD62P)CD15-E-selectin(CD62E)、P-selectin(CD62P)淋巴细胞归位受体-血管内皮细胞地址素:L-selectin(CD62L)-PNAdCLA-E-selectin(CD62E)LFA-1(CD11a/CD18)-ICAM-1(CD54)、ICAM-2(CD102)VLA-4(CD49d/CD29)-VCAM-1(CD106)CD44-MAdLPAM-2(CD49d/β7)-MAd、VCAM-1(CD106)白细胞粘附分子-细胞外基质:VLA-1(CD49a/CD29)-CA、LMVLA-2(CD49b/CD29)-CA、LMVLA-3(CD49c/CD29)-FN、LM、CAVLA-4(CD49d/CD29)-FNVLA-5(CD49e/CD29)-FNVLA-6(CD49f/CD29)-LMα7β1(-/CD29)-LMVNR-β1(CD51/CD29)-VN、FNMac-1(CD11b/CD18)-FBP150,95(CD11c/CD18)-FBGPⅡbⅢa(CD41/CD61)-FB、FN、vWF、TSPVNR(CD51/CD61)-VN、FB、vWF、FN、CA、TSPα6β4(CD49f/CD104)-LMVNR-β5(CD51/-)-VNCD51/ β6-FNCD49d/β7-FNGPIb-α/IX(CD42b/CD42a)-vWF免疫细胞间相互识别:CD22-CD45ROCD2-LFA-3(CD58)CD4-MHCⅡ类分子CD5-CD72CD 8-MHCⅠ类分子LFA-1(CD11a/CD18)-ICAM-1(CD54)、ICAM-2(CD102)CD28-B7/BB1(CD80)CD27-CD70参与白细胞激活T细胞:CD2(T细胞旁路激活途径)、CD3(信号转导)、CD4、CD5、CD8、CD28、 CD43、CD44、VLA-4(CD49d/CD29)、CDw90B细胞:CD19(抑制G→C1,抑制Ig分泌)、CD20(抑制细胞周期)、CD21(活化B细 胞)、CD22(Ca2+升高,促进G→G1)、CD23(B细胞分化)、CD40(B细胞生 长)、CD72、CD73(G→G1)、CD80(B细胞活化)髓样细胞:CD14(髓样细胞氧化爆发?)、CDw65(中性粒细胞活化)NK:CD16(NK活化)、CD2、CD3非谱系:CD69(活化诱导分子,AIM)与细胞膜表面酶有关CD10(中性肽链内切酶)、CD13(氨肽酶)、CD26(二肽酰肽酶Ⅳ)、CD45(酪氨酸磷酸酯酶)、 CD73(5`核苷酸外切酶)、CDw75(具有酶活性)与病毒受体有关CD21(EB病毒R)、CD4(HIVR)、CD54(鼻病毒R)、CD46(麻疹病毒受体) | |
一、与T细胞识别、粘附、活化有关的CD分子
T细胞是一类重要的免疫活性细胞,除直接介导细胞免疫功能外,对机体免疫应答的调节起关键作用。T淋巴细胞本身的识别活化及效应功能的发挥,不仅与外来抗原、丝裂原和多种细胞因子密切相关,而且有赖于T细胞相互之间、T细胞与抗原提呈细胞(APC)之间以及T细胞与靶细胞之间的直接接触。T淋巴细胞识别抗原的受体是T细胞受体(t cell receptor,TCR)与CD3所组成的复合物(TCR/CD3)。在识别过程中还有赖于抗原非特异性的其它细胞表面分子的辅助,这些辅助分子(accessory molecules)主要包括CD4、CD8,MHC Ⅰ类分子、Ⅱ类分子,LFA-1(CD11a/CD18)、CD49d、e、f/CD29(VLA-4、VLA-5、VLA-6)、CD28、CD44、CD45、ICAM-1(CD54),LFA-2(CD2)和LFA-3(CD58)等。
有关MHCⅠ类、Ⅱ类分子的结构和功能在第六章“MHC及其临床应用”中讨论。VLA-4、VLA-5、LFA-6、LFA-1、ICAM-1、CD44见第二章“粘附分子”。有关CD45在第八章“免疫球蛋白超家族”中阐述。
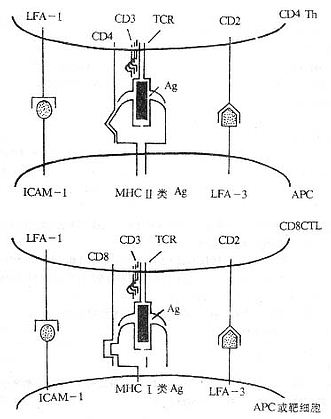
图1-1 参与T细胞对靶细胞识别的分子(模式图)
T细胞表面的辅助分子有以下特点:
(1)存在于T细胞上的辅助分子可特异地与存在于APC或靶细胞上的某些分子(配体)相结合,如LFA-1和CD2可分别与ICAM-1和LFA-3结合。
(2)辅助分子本身不具有多态性,在一个物种所有个体的所有T细胞的某一种辅助分子的结构基本上是相同的。
(3)辅助分子可加强T细胞与APC或靶细胞结合的程度。
(4)许多辅助分子具有转导信号的功能,如CD2、CD4和CD8等分子。
(5)有些辅助分子如CD2、CD4、CD8、CD28、Thy-1等其编码的基因属于Ig基因超家族;有些辅助分子如LFA-1、VLA-4、VLA-5和VLA-6等编码的基因属于integrin 基因超家族。
(6)T细胞膜表面辅助分子作为膜表面重要的标记已被应用于临床的诊断和治疗。
(7)细胞因子可调节辅助分子的表达,从而改变细胞间粘附的能力,这是细胞因子免疫调节作用的一个重要方面。
(一)T细胞受体
T细胞受体(T cell receptor,TCR或Ti)是T淋巴细胞表面识别外来抗原与自身MHc Ⅰ类抗原(或Ⅱ类抗原)复合物的受体,在同种异体移植中TCR也识别单独的非已的MHC抗原。目前已经证实,TCR在细胞表面与CD3密切结合在一起组成TCR/CD3复合物,TCR识别抗原后刺激信号是通过CD3分子传递的。
1.T细胞受体的类型和结构 TCR中的多肽链是异质性的。根据抗原结构和编码基因不同,已发现有α、β、γ和δ四种多肽链。关于TCR多肽链的结构大多是从分析TCR多肽链cDNA或基因组克隆(genomicclones)而来,编码TCR多肽链的基因属于免疫球蛋白基因超家族成员。成熟TCR肽链分子量在40~60kDa之间。根据TCR中异源双体的组成的不同,TCR可分为以TCRαβ和TCRγδ两种类型。
(1)TCRαβ:CD阳性TCRαβT细胞可识别非已MHCⅡ类抗原(同种异体抗原)或自身MHCⅡ类抗原与加工后抗原的复合物CD8阳性TCr αβT细胞则可识非已MHCⅠ类抗原或自身MHC Ⅰ类抗原与加工后抗原的复合物TCRα链分子量40~50kDa的酸性糖蛋白,β链40~50kDa不带电或碱性糖蛋白。α和β链各由一个可变区(V区)和一个恒定区(C区)组成,与Ig的V区和C区大小相似,属于免疫球蛋白超家族成员。TCRα、β链的V区约含102到109个氨基酸,在V区部分由两个半胱氨酸形成链内二硫键,组成约含50~60氨基酸残基的环肽,这与IgV区结构和功能相似,是特异性识别外来抗原的结构域。TCRα、β链的C区约含138到179个氨基酸,每个C区形成由链内二硫键连接的环肽。α、β链在连接肽(connectingpeptide)形成链间二硫键。穿膜区约由20~24氨基酸组成,α链穿膜区含有带正电的1个赖氨酸和1个精氨酸残基,β链穿膜区含有1个带正电的赖氨酸残基,这些带正电的氨基酸与CD3γ、δ和ε链穿膜区带负电的谷氨酸和/或天冬氨酸形成盐桥,稳定TCR/CD3复合物结构,并与CD3传递信息有关。α、β链胞浆部分只有5~12氨基酸长的尾部(图1-2)。
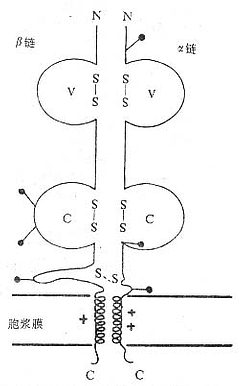
图1-2 TCRαβ异源双体模
(2)TCR γδ:TCRγ和δ链各包括一个Ig样的V区和C区、连接肽、疏水的穿膜区以及一个短的胞浆区尾部,在连接肽区可形成链间的二硫键。γ和δ链的穿膜区各含有1个带正电的赖氨酸,此外δ链还有1个带正电的精氨酸,这些带正电的氨基酸与CD3γ、δ和ε链穿膜区带负电的天冬氨酸或谷氨酸形成盐桥。在氨基酸水平上分析,TCRγ链与β链同源性较高,而TCRδ链与α链同源性较高。在人类TCRγδ有二硫键相连和非共价相连两种形式,而在小鼠只发现二硫键相连的TCRγδ形式。人γ链分子量为36~55kKa,δ链为40~60kDa,γ、δ链的分子量大小取决于多肽骨架的长度和糖基化的程度。
有关TCRα、β、γ、δ链基因的结构和重排见第三章“免疫球蛋白超家族”
2.两种类型TCR T细胞的比较 TCRαβ与TCRγδ不仅组成受体多肽链的结构不同,而且具有这两种类型受体T细胞的分布、表型、发育以及功能也有差别(表1-5)。
表1-5 TCRαβ与TCRγδ细胞特性的比较
| 特性 | TCRαβ | TCRγδ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 分 布 | PBL | 60~70% | 0.5~15% | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 其它部分 | 小鼠树突状表皮细胞(DEC)、小鼠粘膜上皮内淋巴细胞(IEL) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 表 型 | CD4+CD8- | 60~65% | 50000/巨噬细胞,80%同源性),至少可分为11个家族(family).人V基因片段约为100个,至少可分为6个家族,每个家族含有2~60个成员不等。V基因片段由2个编码区(codingregions)组成:第一个编码区编码大部分信号序列;第二个编码区编码信号序列羧基端侧的4个氨基酸残基和可变区约98个氨基酸残基,包括互补决定区1和2(complementarity determining region 1和2,CDR1和CDR2)。 (2)重链D基因片段:D(persity)是指多样性。DH基因片段仅存在于重链基因中而不存在于轻链基因。D基因片段编码重链V区大部分CDR3。小鼠DH共有12个片段,位于VH和JH基因片段之间,大部分DH片段较为集中,约占60~80kb,但靠上游的DH可能位于VH区域内,最后一个DH片段与JH基因5端相距约0.7kb。人类DH片段可能有10~20个左右。 (3)重链的J基因片段:J(joining)指连接,是连接V和C基因片段。JH编码约15~17个氨基酸残基,包括重链V区CDR3除DH编码外的其余部分和第4骨架区。小鼠JH基因片段有4个,与Cμ相距约6.5kb。人有9个JH,其中6个是有功能的JH基因片段。 V、D、J基因片段经重组连接在一起,组成2个外显子,一个外显子编码信号序列的大部分,另一个外显子编码信号序列的其余部分和重链可变区。 小鼠和人在胚系中Ig基因的结构见图3-4和图3-5。 2.重链可变区基因的移位 在重链基因重排开始时,二条染色体上都发生D基因片段移位到J基因片段而发生D-J基因连接。在此以后,只有其中一条染色体上的V基因片段与D-J基因片段连接。VH基因片段5端含有启动子(promoter),JH和Cμ基因片段之间的内含子中含有转录增强子(transcriptinalenhancer)。如果一条染色体VH基因与D-J基因重排无效(non-productive),另一条染色体的VH基因片段开始发生移位,与D-J基因片段连接。 某些与Ig基因片段重排有关的特殊序列称为识别序列(recognition sequences),位于V基因片段的3端与J基因片段的5端之间以及D基因片段的两侧。V基因片段3端、J基因片段5端以及D基因片段的两侧也是DNA重排识别信号所在区域,这些识别信号包括三部分:(1)高度保守的回文结构的七聚体(palindromic heptamer);(2)较少保守、富含A/T的九聚体(nonamer);(3)七聚体和九聚体之间不保守的间隔序列(spacer sequence),含有12±1碱基对或23±1碱基对。根据12/23碱基对间隔规则(或称1圈/2圈定律),两个基因片段的重组仅发生在两个基因片段之间:各有一个12个碱基对片段和一个23个碱基对片段的结构(图3-6)。
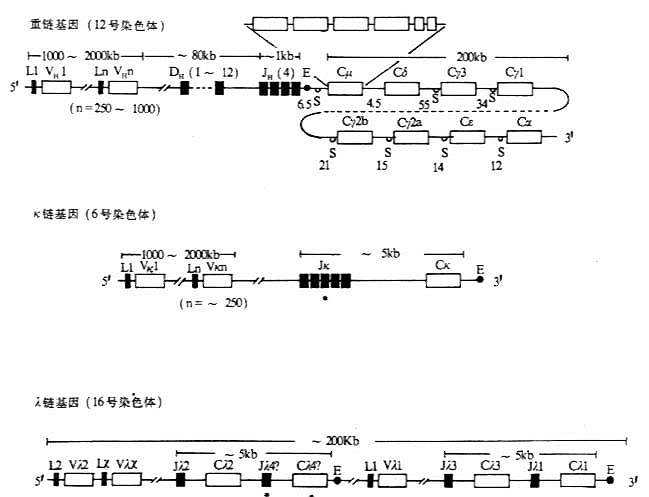
图3-4 小鼠Ig基因结构 注:(1)L:先导序列基因片段 V:可变区基因片段 D:D基因片段 J:J基因片段 C:恒定区基因片段 E:转录增强子 S:转换区 *:假基因 (2)内含子区域所标数字表示DNA长度(kb)。 (3)每个CH基因用一个方框表示,实际上包括几个外显子,如Cμ含有6个外显子。 (4)λ基因增强子位于Cλ4和Cλ1基因片段的下游。
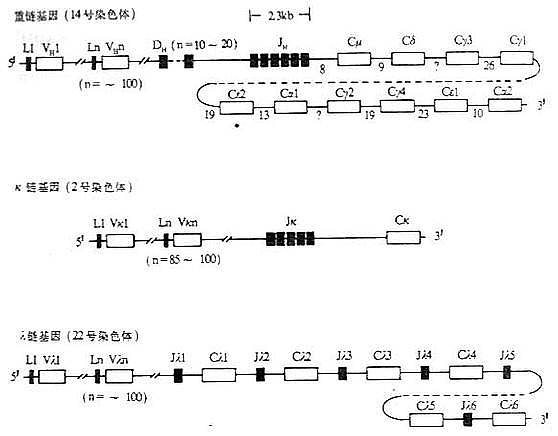
图3-5 人Ig基因结构 注:(1)L:先导序列基因片段 V:可变区基因片段 D:D基因片段 J:J基因片段 C:恒定区基因片 *:假基因 (2)内含子区域所标数字表示DNA长度(kb) (3)每个CH基因用一个方框表示,实际上包括几个外显子 参与V/(D)/J基因重组过程的酶称为V/(D)/J重组酶(recombinase),有关于执行识别、切割和重新连接基因片段重组酶的纯化和鉴定工作还刚开始。重组酶实际上包括重组过程中多种酶的活性。最近在前B细胞(pre-b cell)中已经鉴定出两种刺激Ig基因重排的基因,称为重组激活基因1(recombinationactivating gene 1,RAG-1)和重组激活基因2(RAG-2),其确切的作用机理还不太清楚。重组酶作用的特点是:(1)淋巴细胞特异性的,非淋巴样细胞如成纤维细胞无重组酶活性,这可能解释了Ig基因的重排仅见于B淋巴细胞。目前一般认为T细胞TCR基因重排中的重组酶与B细胞中重组酶相同或相似。(2)重组酶发挥其功能仅限于B细胞发育早期,未成熟B细胞如前B细胞(pre-b cell)细胞系重组酶活性很高,但抗体生成细胞或骨髓瘤细胞无明显重组酶活性,因此时B细胞已经分泌某一特异性抗体,不再发生重排其它的Ig基因,因此也不会改变原先所产生抗体的特异性。转换重组酶(switch recom-binase)可能与VDJ重组酶(VDJ recombinase)相似,但缺乏七聚体/九聚体(heptamer/nonamer)识别蛋白。重组酶功能异常可导致机体不能产生Ig和TCR,很可能与重症联合免疫缺陷(severe combinedimmunodeficiency,SCID)的发生有关。例如SCID小鼠16号染色体着丝点末端存在一个scid基因,为单基因常染色体遗传基因。scid基因纯合将影响DNA重组酶的识别功能,在TCR或BCR基因片段重排时不能识别正确的位点,使T细胞、B细胞在淋巴干细胞发育早期即夭折,导致重症联合免疫缺陷。
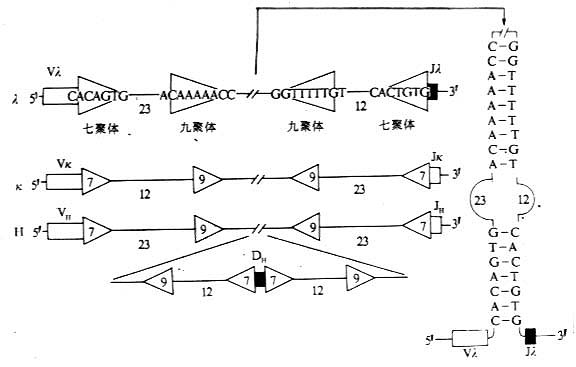
图3-6 参与Ig基因重排组酶识别的DNA序列 (二)Ig重链恒定区(C区)基因 1.重链C基因片段 重链恒定区基因由多个外显子组成,位于J基因片段的下游,至少相隔1.3kb。每1个外显子编码1个结构域(domain),铰链区(hinge region)是由单独的外显子所编码,但α重链的铰链区是由CH2外显子的5端所编码。大多分泌的Ig重链羧基端片段或称尾端“tail piece”是由最后一个CH外显子的3端所编码,而δ链的“tailpiece”是由一个单独的外显子所编码。小鼠CH基因约占2000kb,其外显子从5端到3排列的顺序是Cμ-Cδ-Cγ3-Cγ1-Cγ2b-Cγ2a-Cε-Cα。人CH基因外显子排列的顺序是Cμ-Cδ-Cγ3-Cγ1-Cε2(pseudo基因)Cα1-Cγ2-Cγ4-Cε1-Cα2。其中基因片段Cγ3-Cγ1-Cε2-Cα1和基因片段Cγ2-Cγ4-Cε1-Cα2可能是一个片段经过一次复制而得,为研究CH基因的起源和进化提供有用的依据。 2.免疫球蛋白类型转换 1964年Nossal等发现B淋巴细胞存在着类型的转换。Ig类型转换(class switch)或称同种型转换(isotype switch)是指一个B淋巴细胞克隆在分化过程中VH基因片段保持不变,而发生CH基因节段的重排、比较CH基因片段重排后基因编码的产物,V区相同而C区不同,即识别抗原特异性不变,而类或亚类发生改变。这种类型转换在无明显诱因下可自发产生。 局部微环境和细胞因子可影响和调节免疫球蛋白类型的转换,如在肠道派伊尔氏结的B细胞V基因片段优先转换到Cα1进行重排,因此主要合成和分泌IgA。在体外向经LPS刺激的小鼠B细胞中加入IL-4,可促进B细胞产生IgG1和IgE,抑制IgG2b产生;低浓度的IL-4主要诱导产生IgG1,高浓度IL-4主要诱导产生IgE。面IFN-γ则诱导小鼠B细胞合成IgG2a,抑制IgE的产生。TGF-β、IL-5和IL-6对IgA的产生具有促进作用(表3-3)。细胞因子调节B细胞Ig类别转换的机理可能是:(1)刺激某些细胞的克隆选择性的增殖,使分泌某特定类、亚类抗体的克隆细胞增加,如IL-5、IL-6促进IgA。产生除通过同种型转换进行调节外,还可选择性促进IgA定向细胞分化增殖为IgA分泌细胞。(2)通过诱导特定位置上两个转换区的重组,诱导B细胞由分泌IgM向某一同种型Ig转换。如高浓度IL-4促进LPS诱导小鼠B细胞产生IgE,主要是使Cε转换区与重组酶的接近(accessibility),通过同种型转换促进IgE的产生。 表3-3 细胞因子对LPS诱导的Ig类和 亚类转换的调节作用(占总Ig%)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||